The cytoskeleton of dendrites forms a uniform gel in the spine neck and head
Why maintaining gel-like networks makes sense as a mechanism for brain preservation

Eberhardt et al. 2022, “A uniform and isotropic cytoskeletal tiling fills dendritic spines”, is another data point that it’s useful to think of neural structures as gels.
This is a paper from the Ellisman lab using serial section EM tomography to study dendritic spines. They were able to show that even after segmenting a representative dendritic spine into the membrane, extracellular space, and organelles, electron-dense regions remain, thought to be actin filaments and/or actin-binding proteins:
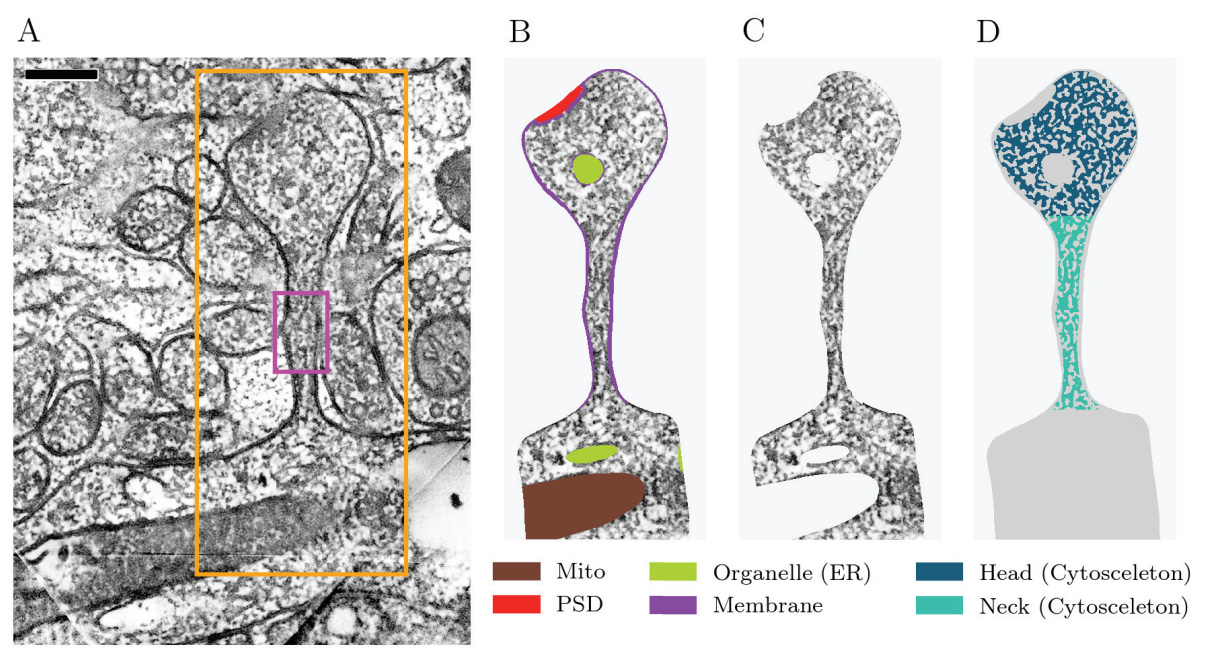
Here is a 3d visualization of just the dendritic spine neck (E), followed by a binarization algorithm (F), a visualization of the topological skeleton of the presumed actin filaments (G), and a reconstruction of the actin filaments from the skeleton (H):

Here are some more of the spines they analyzed. The top ones are from Purkinje cells, while the bottom ones are from hippocampal pyramidal cells:
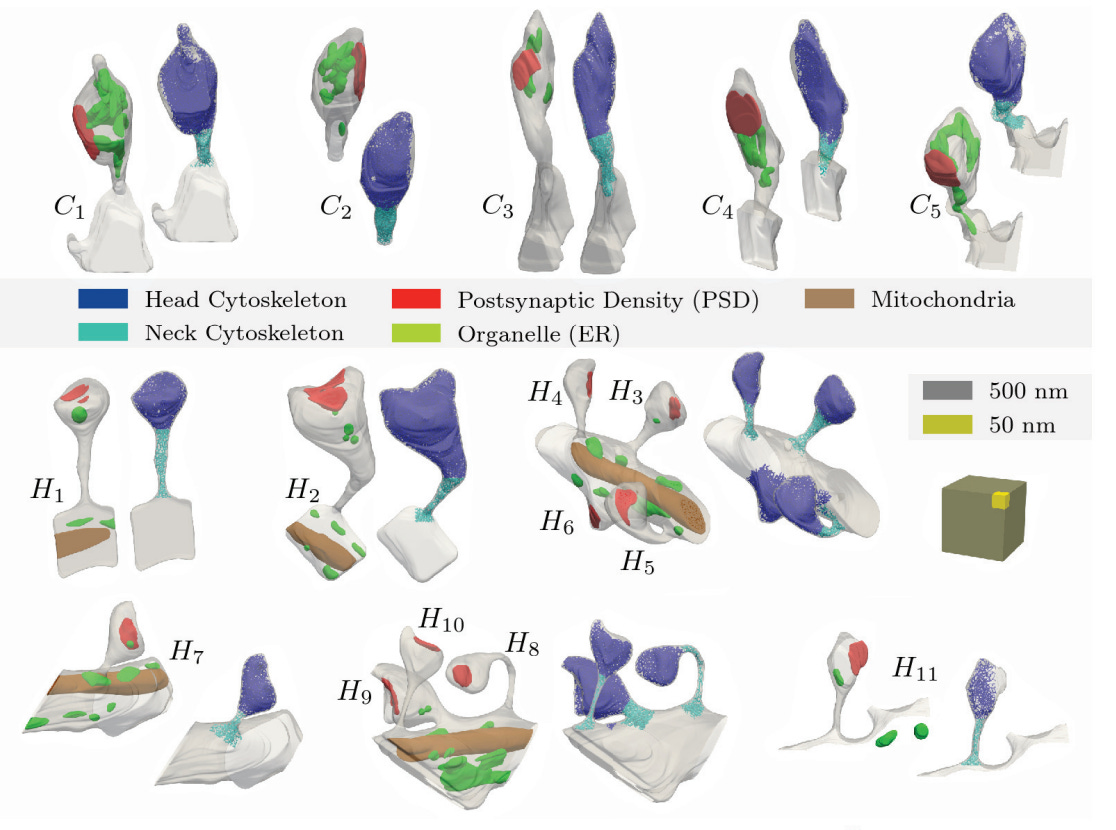
Here are some of the key points from their discussion:
“The cytoskeleton in spines must serve two (possibly) opposing purposes: confer mechanical stability and permit the passage of molecules through the gel.”
“Glutaraldehyde fixation prior to high-pressure fixation and freeze substitution minimized the extraction of soluble cytoplasmic proteins, which meant that these were also stained. The ultrastructure we extracted was, therefore, not exclusively composed of actin and actin-binding proteins. A quantitative study comparing cryoEM to aldehyde fixation finds that hippocampal spine neck diameters are 50% larger after aldehyde fixation (Tamada et al. 2020), which indicates a possible role of spurious cross-links arising from cytoplasmic proteins in expanding the actin network. In our hands, however, the measured spine neck diameters were comparable to the cryoEM measurements of Tamada et al."
“We tracked filaments traversing both the actin branches and numerous cross-links. The reconstructed mesh, therefore, extended beyond F-actin and its branches, with the cross-linking proteins creating a uniform actin-polymer gel throughout all spine sub-domains. Despite differences in size between cerebellar and hippocampal spines, the statistical properties of the cytoskeletal ultrastructure were quite similar in both cell types.”
“Most differences in statistical measures of the internal structure between spine necks and heads could be explained by differences in the surface-to-volume ratio, with the exception of a higher density of putative actin and nodes in spine heads (Fig. 2E, 4D). These differences were minor, however, and smaller in magnitude than the standard deviation across samples. Taken together, these results provide evidence against the traditional view that spine heads and spine necks possess distinct actin architectures (Halpain, 2000, Hotulainen et al., 2009, Tada and Sheng, 2006). The in-vitro studies of Korobova and Svitkina (2010) have already shown that the actin-cytoskeleton undergoes a more gradual and less drastic change in its architecture from the spine head to the neck. Our in-situ data go a step further and demonstrate that the cytoskeleton is a uniform gel down to the nanometer scale.”
What are some of the implications of this work for brain preservation?
Humans think and reason largely through narratives, and in the case of biology, mechanisms. For better or worse (often worse, I think), it’s not enough for us to have empirical evidence that structure is likely maintained by a particular preservation procedure. People need to know why.
The mechanism of cryopreservation, i.e. slowing biological time, is widely known. On the other hand, the mechanism of aldehyde fixation is not usually expanded upon in great depth. Much is made of “crosslinking” of molecules, but it isn’t entirely clear why that should maintain structures over time.
I think that understanding the aldehyde fixation story requires us to understand the structure of cells prior to fixation. Are cells solid or liquid? If they are a liquid, why don’t they just fall apart?
I think that gels — a form of matter that has both solid-like and liquid-like properties — are the best explanation for this. The cytoskeleton, and other gel-like structures inside and outside of cells, maintain neural structures by establishing a gel-like mesh of molecules, actin chief among them.
After death, these gels start to break down due to proteolysis and diffusion, and eventually, they completely liquefy. By that point, the structural information is lost. An obvious mechanism of structural brain preservation, then, is to stop or at least dramatically slow the breakdown of gel-like networks so that the morphological information can be maintained. And that seems to be what crosslinking aldehydes accomplish.
This paper shows us that dendritic spines have cytoskeletons that are uniform gels throughout all sub-domains, which means they should be preserved by a fixation procedure that stabilizes gels.
While empirical data has already suggested this for some types of dendritic spines, corroboration of it, including showing that this is a widespread phenomenon across neuronal types and sub-domains, is good corroboration that the existing empirical data will generalize.
Since dendritic spines are one of the key parts of the brain that have been associated with memory, it is very helpful to be able to reason about what will happen to these structures during a brain preservation procedure.
If restoring dendritic spines from fixative-based brain preservation ever becomes possible, it would require nanotechnology to physically remove the aldehyde crosslinks, or scanning and emulation technology to remove them in silico. Among many other requirements! This is just with regard to the aldehyde crosslink component.
Reference
Eberhardt F, Bushong EA, Phan S, et al. A uniform and isotropic cytoskeletal tiling fills dendritic spines. eNeuro. Published online October 10, 2022:ENEURO.0342-22.2022.